Скачать статью в PDF формате
English version
УДК 595.598.132
Поступила в редакцию: 28.02.2017
Принята в печать: 01.07.2017
Для цитирования:
Акрамова Ф.Д., Медетов М.Ж., Шакарбаев У.А., Азимов Д.А., Сапаров К.А. Прямокрылые (Orthoptera: Acrididae) Узбекистана – промежуточные хозяева филяриат паразитов птиц // Российский паразитологический журнал. - М. - 2017. - Вып. . - С. .
For citation:
Akramova F.D., Medetov M.J., Shakarbaev U.A., Azimov D.A., Saparov K.A. Orthoptera (Orthoptera: Acrididae) of Uzbekistan is intermediate hosts filyariat of parasites of birds // Russian Journal of Parasitology, 2017, V. , Iss. , pp. .
Прямокрылые (Orthoptera: Acrididae) Узбекистана – промежуточные хозяева филяриат паразитов птиц.
Акрамова Ф.Д. 1, Медетов М.Ж. 1, Шакарбаев У.А. 1, Азимов Д.А. 1, Сапаров К.А.2
1 – Институт генофонда растителного и животного мира АН РУз, ushakarbaev@mail.ru
2 – Ташкентсклй государственный педагогический университет
Реферат
Цель исследования – исследование биологии и жизненных циклов некоторых видов филяриат – паразитов птиц в условиях Узбекистана.
Материалы и методы. Собрано и исследовано более 15 тыс. экз. прямокрылых из сем. Acrididae по общепринятым методам энтомологии и паразитологии. Проведены опыты по заражению птиц личинками филярий обнаруженных у прямокрылых.
Результаты и обсуждение. У 6 видов прямокрылых сем. Acrididae обнаружены инвазионные личинки двух видов нематод Aprocta cylindrica и Diplotriaena isabellina в ряде регионов Узбекистана. Экспериментально установлено участие саранчовых в качестве новых промежуточных хозяев указанных видов нематод.
Ключевые слова: нематоды, филяриаты, прямокрылые, саранчовые, Узбекистан.
Введение
Филяриаты широко представлены среди птиц во многих регионах мира (Baruš et al., 1978; Anderson, 2000; Saparov et al., 2013). Они паразитируют у птиц различных экологических групп. Для ряда видов Aprocta и Diplotriaena в качестве промежуточных хозяев установлены саранчовые (Anderson, 1962, 2000; Quentin et al., 1976; Кабилов, 1983). Этими данными ограничиваются сведения о биологии представителей родов Aprocta Linstow, 1883 и Diplotriaena Henry et Ozoux, 1909, которые включают около 110 видов мировой фауны, паразитирующих в воздухоносных мешках, легких и полости тела птиц различных отрядов, преимущественно, воробьинообразных.
Целью настоящей работы является исследование жизненных циклов нематод A. cylindrica и D. isabellina – паразитов птиц в условиях Узбекистана.
Материал и методы
Материалом настоящего исследования послужили собственные сборы саранчовых Acrididae отряда прямокрылых в местах концентрации птиц в районах Ташкентской, Сырдарьинской, Джизакской, Кашкадарьинской, Бухарской областей и Республики Каракалпакстан. Всего в разные сезоны 2010-2016 гг. (весна, лето, осень) собрано и исследовано 15342 экз. саранчовых по общепринятой методике (Nelson, 1960; Кабилов, 1983; Бекузин и др., 1993; Лачининский и др., 2002; Хамраев и др., 2008). Обнаруженные личинки нематод фиксировали в 1.5-2.0 % формалине. Основную часть личинок помещали на предметное стекло в капле физиологического раствора и приводили их в неподвижное состояние легким подогреванием над спиртовкой, затем накрывали покровным стеклом и исследовали под микроскопом. Птиц (стерильных) заражали инвазионными личинками, выделенными из саранчи. В эксперименте использовали 4 ос. птенцов домового воробья, по 3 птенцов кеклика, перепела и майны. Опытные птицы были заражены по 33-45 экз. личинок. Для контроля использовали птенцов по 1 ос. каждого вида. Исследования проводили современными приборами: микроскоп инвентированный СК-2-TR (Olуmpus, Japan), бинокуляр ML-200 (Olуmpus, Japan).
Результаты и обсуждение
Мы в процессе сбора биологического материала в районах ряда областей и Республики Каракалпакстан у представителей саранчовых (Acrididae) находили личинок нематод в различных стадиях развития. Среди них были инвазионные личинки третьей стадии. Естественная зараженность саранчовых - Calliptomus turanicus (Tarbinsky, 1930), Dociostaurus kraussi (Ingenizkij, 1897), Locusta migratoria (Linnalus, 1758), колебалась от 2.2 до 3.8%, при интенсивности инвазии 3-11 экз. Личинки свободно находились в их жировом теле. Тело инвазионных личинок цилиндрическое, 0.462 - 0.508 мм длины, 0.040 - 0.046 мм ширины, к переднему и заднему концам суживается. Нервное кольцо находится на расстоянии 0.052-0.056 мм и экскреторное отверстие 0.072-0.078 мм от головного конца. Пищевод длиной 0.410-0.520 мм. Кутикула поперечно исчерчена. Тело личинки, особенно кишечник, обильно заполнен зернистыми тельцами (рис.1).
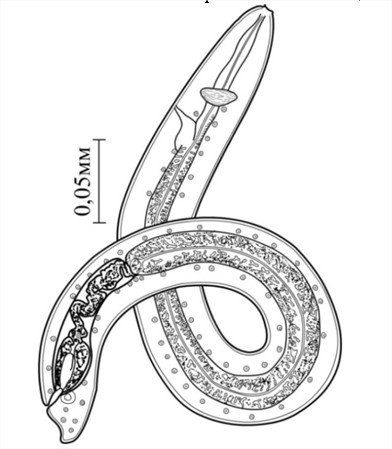
Рис.1. Aprocta cylindrica Linstow, 1883, Larvae от саранчи Calliptomus turanicus Tarb: инвазионная личинка III-стадии.
По морфологическим признакам изучаемые личинки очень сходны с личинками A. cylindrica, описанными Quentin et al. (1976) от саранчи – Locusta migratoria.
Инвазионные личинки от каждого вида саранчи были скормлены подопытным птенцам домового воробья Passer domesticus (Linnaeus, 1758). Экспериментальное заражение птенцов проводили в июле 2015 года. Птенцы вскрывались через 25, 35, 45 дней после заражения с целью установления развития инвазионных личинок до зрелых форм нематод в организме дефинитивного хозяина (табл. 1).
Таблица 2
Экспериментальное заражение птиц инвазионными личинками Aprocta cylindrica Linstow, 1883, выделенных от саранчовых Узбекистана
|
Птенцы |
Calliptamus turanicus |
Dociostaurus kraussii |
Locusta migratoria |
Сроки иссл. (дни) после заражения |
|
Число личинок |
||||
|
Passer dom.: |
|
|
|
|
|
№1 |
34 |
- |
- |
25 |
|
№2 |
- |
36 |
- |
35 |
|
№3 |
- |
- |
37 |
45 |
|
№4 |
контроль |
45 |
||
Как показали результаты исследования через 25 - 35 дней после заражения в носовой полости у подопытных птенцов №1, 2 были обнаружены развивающиеся ювенальные формы нематод самцов и самок, у которых хорошо заметны половые органы. На 45 день у птенца №3, также в носовой полости найдены зрелые нематоды: 5 самцов и 11 самок. У контрольных птиц (№4) нематоды не были обнаружены. Морфологические параметры самцов и самок находились в полном соответствии с признаками зрелых червей A. cylindrica, обнаруженных нами у ряда видов воробьинообразных и данными литературы (Сонин, 1966; Боргаренко, 1990). При этом в носовой полости также было обнаружено значительное число отложенных яиц, с толстой скорлупой. Размеры яиц колебались 0.042 - 0.056 мм длины и 0.028 - 0.036 мм ширины.
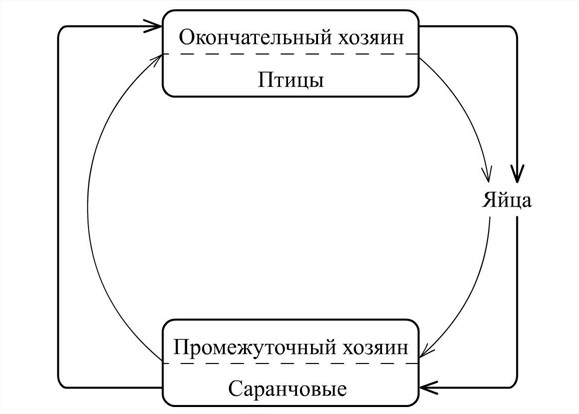
Рис.2. Схема цикла развития Aprocta cylindrica Linstow, 1883.
Таким образом, в процессе развития личинок в организме позвоночного хозяина происходит дальнейшее увеличение длины и ширины тела, причем темпы роста на этой стадии различны у самцов и самок. Наблюдается интенсивное развитие половой системы у обоих полов. На 45-й день после заражения были полностью сформированы, практически все органы филярий рассматриваемого вида и, вероятно, они приступают к спариванию. Об этом могут свидетельствовать отложенные яйца, обнаруживаемые в просвете и тканях носовой полости зараженных птиц.
У подопытных птиц на 45 – й день после скармливания им инвазионных личинок нематоды достигают половой зрелости (рис. 2).
При исследовании саранчовых Узбекистана нами также выявлены личинки D. isabellina в различных стадиях развития у следующих видов: Bryodema tuberculatum (Fabricius,1775), Aiolopus oxianus (Uvarov, 1926), Melanoplus frigidus (Boheman, 1846) и Calliptamus italicus (Linnaeus, 1758). Интенсивность инвазии составила у отдельных видов саранчи от 1 до 7 экз. Инвазионные личинки были обнаружены в жировом теле, окруженные тонкостенной, прозрачной оболочкой (рис. 3). Экстенсивность инвазии саранчовых составила в пределах 2.1-4.0 %.
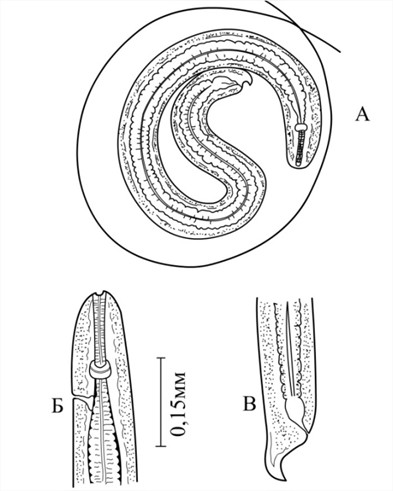
Рис. 3. Diplotriaena isabellina Koroliowa, 1926, Larvae от Calliptamus italicus (L.), инвазионная личинка III-стадии: А-общий вид личинки в цисте, Б-передний конец,
В-хвостовой конец.
Тело инвазионной личинки цилиндрическое. Длина 0.528 – 0.641 мм, при максимальной ширине 0.042 – 0.048 мм. К переднему и заднему концам резко суживается. От переднего конца тела до нервного кольца 0.057 – 0.060 мм, до экскреторного отверстия 0.076 – 0.080 мм. Пищевод состоит из двух отделов. Мышечный отдел пищевода – 0.074 – 0.076 мм, железистый – 0.460 – 0.464 мм, длины. Вершина хвоста удалена от анального отверстия на 0.038 – 0.040 мм. Кутикула с тонкой поперечной исчерченностью. Тело личинки заполнено зернистыми тельцами (рис. 3). По морфометрическим признакам изучаемые личинки очень сходны личинкам D. isabellina (Кабилов, 1983).
После тщательного изучения инвазионных личинок, мы проводили серии опытов по заражению ими птенцов перепела, кеклика и майны (табл. 2).
Таблица 2
Результаты экспериментального заражения птиц инвазионными личинками Diplotriaena isabellina
|
Вид птицы |
Число личинок, экз. |
Дни исслед. после заражения |
||||
|
Coturnix coturnix (перепел) |
№1 |
33 |
35 |
|||
|
№2 |
36 |
40 |
||||
|
№3 |
45 |
50 |
||||
|
№4 |
Контроль |
55 |
||||
|
Alectoris chucar (кеклик) |
№1 |
35 |
40 |
|||
|
№2 |
45 |
55 |
||||
|
№3 |
50 |
65 |
||||
|
№4 |
Контроль |
65 |
||||
|
Acridotheres tristis (майна) |
№1 |
30 |
45 |
|||
|
№2 |
33 |
55 |
||||
|
№3 |
35 |
65 |
||||
|
№4 |
Контроль |
65 |
||||
При вскрытии перепела №1 через 35 дней после заражения, кеклика №1 – 40 и майны №1 – 45 дней находили ювенальные формы самцов и самок филярий. Нематоды были обнаружены в просвете легких и воздухоносных мешков. Через 45 – 55 дней нематоды отмечены у остальных подопытных птиц, преимущественно, в просвете воздухоносного мешка. Органы самцов и самок достаточно четко оформлены, но они еще не достигли зрелого состояния. Среди общей массы нематод встречались единичные экземпляры, как самцов, так и самок, размеры которых соответствовали зрелым паразитам. Длина самцов составляла 19-20 мм, самок – 28-35 мм. Как показали вскрытия подопытных птиц (майны №3 и кеклика №3), большинство обнаруженных паразитов были зрелыми. В воздухоносном мешке майны обнаружены 8♂ и 13♀, а у кеклика – 7♂ и 11♀.
В целом, биологический цикл развития, рассматриваемой нематоды, соответствует известным данным (Anderson, 1962) с той лишь разницей, что в качестве промежуточных хозяев D. isabellina мы отмечаем три новых вида саранчовых - Aiolopus oxianus, Calliptamus italicus и Melanoplus frigidis. Сроки развития паразита в организме дефинитивных хозяев в наших опытах происходили значительно быстрее, чем в экспериментах Anderson (1962). Вероятно, это связано с условиями опытов и степенью восприимчивости различных видов птиц. По данной схеме биологический цикл, вероятно, будет осуществляться у остальных видов рода, которые локализуются в органах, сообщающихся с внешней средой (воздухоносные мешки, орбитальные полости, полости носа). Подтверждением этой точки зрения могут служить, данные, полученные рядом авторов (Bain, Vaucher, 1973; Cawthorn, Anderson, 1980), которые зарегистрировали в качестве промежуточных хозяев для некоторых видов рода Diplotriaena - D. agelaeus (Walton, 1927), D. tricuspis (Fedtschenko, 1874), D. tridens (Molin, 1858), D. bargusinica (Skrjabin, 1927) представителей саранчовых (Camnula pellucidа, Melanoplus bilituratus, M. fusciatus, M. sanguinipes, Locusta migratoria, Schistocerca gregaria).
Обобщая данные наших экспериментальных исследований по жизненным циклам A. cylindrica и D. isabellina и литературы (Anderson, 1962, 2000; Bain, Vaucher, 1973; Quentin et al., 1976; Cawthorn, Anderson, 1980; Кабилов, 1983) можно отметить, что эти нематоды паразитируют в воздухоносных мешках многих видов птиц различных экологических групп, главным образом, воробьинообразных. Самки откладывают яйца, содержащие личинку. Яйца, через дыхательную трубку, попадают в пищеварительный тракт птицы и с испражнениями попадают во внешнюю среду. Промежуточные хозяева (саранчовые) заглатывают яйца с пищей. Личинки, вышедшие из яиц мигрируют и проникают в гемоцель и жировое тело. Развивается до инвазионной стадии, претерпевая двукратную линьку. Заражение птицы происходит при поедании ею саранчовых, содержащих инвазионных личинок.
В процессе сбора саранчовых из природных биотопов у некоторых особей саранчи туранской, атбасарки, марокской, обитающих в низовьях Амударьи (территория Кунградского и Муйнакского районов Каракалпакстана, июль-август 2015-2016 гг.) нами замечано неадекватное их поведение. Отдельные особи находились в состоянии малой активности на ветках растений или почве. На фоне слабой активности, наблюдалось ослаблении реакции избегания (бегства). Можно было их легко ловить руками. Между тем, основные популяции были весьма активными и с трудом их ловили с помощью сачков. Особи с малой активностью указанных видов саранчи, как показали исследования, оказались инвазированными личинками изучаемых нематод. Инвазированные особи саранчи, как правило, становились легкой добычей птиц и других хишников. Этот феномен требует дополнительных исследований. Здесь лишь стоит отметить, что у интенсивно зараженных особей саранчи реакция избегания может частично или полностью подавляться, хотя другие реакции например, на прикосновение сохраняются.
В ряде случаев при паразитированнии гельминтов наряду с ослаблением организма животного наблюдаются специфические ненормальные изменения в поведении (Алексеев, 1993; Кулмаматов и др., 1994). Отклонение от нормального стереотипа поведения приводит организм к особому положению, выделяя его из общей массы популяции и тем самым делает их более доступной жертвой для хищников или для других животных. Такое состояние, вероятно, способствует попаданию зараженных прямокрылых к оканчательному хозяину.
Указанные виды саранчовые относятся к широко распространенным вредителям растений, встречаются во всех исследованных областях Узбекистана и Республики Каракалпакстан (Хамраев и др., 2008) с одной стороны, и оказались промежуточными хозяевами некоторых видов филяриат – паразитов диких и охотничье – промысловых птиц – с другой.
Таким образом, можно утверждать, что прямокрылые – промежуточные хозяева играют важную роль в циркуляции исследуемых видов нематод, так как они являются обитателями травянистого яруса и могут служить обьектом питания птиц различных экологических групп – окончателных хозяев.
Работа выполнена рамках проекта ФЗМВ-2016-0911194528.
Литература
1. Алексеев А.Н. Система клещ возбудитель и ее эмерджентные свойства. Санкт-Петербург, 1993. -204 с.
2. Боргаренко Л.Ф. Гельминты птиц Таджикистана. Нематоды. – Душанбе: Дониш, 1990. Кн.3. - 260 с.
3. Бекузин А.А., Азимов Д.А., Давлетшина А.Г., Кадырова М.К. Насекомые Узбекистана. – Ташкент: Фан, 1993. – 340 с.
4. Кабилов Т.К. Гельминты позвоночных животных Узбекистана, развивающиеся с участием насекомых. – Ташкент: Фан, 1983. – 128 с.
5. Кулмаматов Э. Н., Исакова Д. Т., Азимов Д. А. Гельминты позвоночных горных экосистем Узбекистана. Ташкент: Фан, 1994. - 152 с.
6. Лачининский А.В., Сергеев М.Г., Чильдебаев М.К., Черняховский М.Е., Локвуд Дж.А., Камбулин В.Е., Гаппаров Ф.А. Саранчовые Казахстана, Средней Азии и сопредельных территорий. – Ларами, 2002. – 387 с.
7. Сонин М.Д. Основы нематодологии. Филяриаты животных и человека и вызываемые ими заболевания. Апроктоидеи. – М., 1966. Т. 17. – 360 с.
8. Сонин М.Д. Основы нематодологии. Филяриаты животных и человека и вызываемые ими заболевания. Диплотриеноидеи.– М., 1968. Т.21. – 390 с.
9. Хамраев А.Ш., Нуржанов А.А., Азимов Д.А., Ульмасбаев Ш.Б. Вредные саранчовые и кузнечиковые насекомые Узбекистана и борьба с ними. – Ташкент, 2008. – 74 с.
10. Anderson R.C. On the development, morphology, and experimental transmission of Diplotriaena bargusinica (Filarioidea: Diplotriaenidae) // Canad.J.Zool., 1962. - №40. – P. 1175-1186.
11. Anderson R.C. On the development, morphology, and experimental transmission of Diplotriaena bargusinica (Filarioidea: Diplotriaenidae) // Canad.J.Zool., 1962. - №40. – P. 1175-1186.
12. Anderson R.K. Nematode parasites of vertebrates: their development and transmission. - New York: CAB, 2000. - 650 p.
13. Baruš V., Sergeeva T.P., Sonin M.D., Ryzhikov K.M. Helminths of fish-eating birds of the Palaearctic Region. I. Nematoda. Academia, Praha, 1978. 318 p.
14. Nelson W.S. The identification of filarial larvae in their vectors. Ind. J. Malariol. 1960. 14(4): -Р. 585–592.
15. Quentin J.C., Troncy P.M. and Barre H. Aprocta cylindrica Linstow, 1883, filaire ovipare parasite d,oiseaux au Tchad. Morphogenese larvaire du nematode // Annales de Parasitologie Humaine et Comparee, 1976. V. 51. – P. 83-93.
16. Saparov K., Akramova F., Azimov D., Golovanov V., Kuchboev A. Biodiversity of filariae (Nematoda: Filariina), parasites of birds in Uzbekistan // J. Zool. - Turkey, 2013. - № 37. – P. 746-752.
© 2017 The Authors. Published by All-Russian Scientific Research Institute of Fundamental and Applied Parasitology of Animals and Plants named after K.I. Skryabin. This is an open access article under the Agreement of 02.07.2014 (Russian Science Citation Index (RSCI)http://elibrary.ru/projects/citation/cit_index.asp) and the Agreement of 12.06.2014 (CABI.org / Human Sciences section: http://www.cabi.org/Uploads/CABI/publishing/fulltext-products/cabi-fulltext-material-from-journals-b...