Скачать статью в PDF формате
English version
ФАУНА, МОРФОЛОГИЯ И СИСТЕМАТИКА ПАРАЗИТОВ
УДК 619:639.3.091
DOI:
Поступила в редакцию 03.04.2017
Принята в печать 23.07.2017
Для цитирования:
Воронина Е. А., Володина В. В., Дьякова С. А., Конькова А. В. Разнообразие паразитов и микроорганизмов у каспийских сельдевых рыб // Российский паразитологический журнал. – М., 2017. ‒ Т.41, Вып. 3. ‒ С.
For citation:
Voronina E. A., Volodina V. V., Dyakova S. A., Konkova A. V. Diversity of parasites and microorganisms of Caspian cereal fish. Russian Journal of Parasitology, 2017, V.41, Iss.3, pp.
РАЗНООБРАЗИЕ ПАРАЗИТОВ И МИКРООРГАНИЗМОВ У КАСПИЙСКИХ СЕЛЬДЕВЫХ РЫБ
Воронина Е. А., Володина В. В., Дьякова С. А., Конькова А. В.
Каспийский научно-исследовательский институт рыбного хозяйства, г. Астрахань, ул. Савушкина 1, e-mail: helen212@yandex.ru
Реферат
Цель исследований – изучить видовой состав паразитов и микроорганизмов, а также определить степень зараженности каспийских сельдевых рыб.
Материалы и методы. Объектами исследований служили каспийский и большеглазый пузанок, долгинская сельдь, выловленные на акватории Северного Каспия в весенний период 2014‒2015 гг. С целью изучения паразитофауны морских сельдевых рыб патологоанатомическому вскрытию подвергнуто 88 экз. долгинской сельди и 36 экз. каспийского пузанка. Для микробиологических исследований отобрано по 15 экз. каспийского и большеглазого пузанков и 15 экз. большеглазого пузанка. Учитывали число мезофильных аэробных и факультативно-анаэробных микроорганизмов, видовой состав бактерий, их факторы патогенности и антибиотикорезистентность. Сбор и обработку материала осуществляли в соответствии с общепринятыми в паразитологии и микробиологии методиками. При видовой идентификации использовали определители.
Результаты и обсуждение. Паразитофауна долгинской сельди, каспийского и большеглазого пузанков представлена 7 видами, относящимися к классам: Monogenoidea – 1, Crustacea – 1, Trematoda – 2, Nematoda – 2, Acanthocephala – 1. Сельдевые рыбы вызывали широко распространенные кишечные трематоды Pseudopentagramma symmetricum. Наиболее разнообразно фауна паразитов представлена у каспийского пузанка. Флуктуации в структуре паразитофауны связаны с условиями среды обитания, трофическими связями и специфичностью паразитов к своим хозяевам. Показателями происходящих процессов трансформации экосистем являются индикаторные виды паразитов и микроорганизмы рыб. В ходе исследований установлено, что внутренние органы и жабры сельдевых рыб были заражены микроорганизмами. Максимальная численность бактерий отмечена в кишечнике и жабрах. Доминирующее положение занимали микроорганизмы сем. Enterobacteriaceae. Кроме бактерий, в печени и селезенке рыб при микроскопировании отпечатков органов выявлены гифы микромицетов. Все обнаруженные паразиты и микроорганизмы характеризовались эпизоотической или эпидемиологической значимостью, при этом, они сосуществовали на уровне бессимптомного носительства, не вызывая развития инвазионных и инфекционных процессов в организме обследованных рыб.
Ключевые слова: морские мигрирующие сельди, паразитофауна, биоиндикаторные организмы, гифы микроскопических грибов, микробиоценоз, зараженность.
Введение
На современном этапе морские мигрирующие сельди (долгинская сельдь, каспийский и большеглазый пузанки) относятся к резервным объектам промысла со стабильным промысловым запасом. Промысловые ресурсы этих видов в водах России на 2013 г. оцениваются в 51,58 тыс. т. В случае увеличения масштабов и интенсивности прибрежного промысла закидными неводами, морские сельди могут быть существенным дополнением к общему объему вылова морских видов рыб на юге России [10].
Эта группа гидробионтов представляет большой интерес не только по своему промысловому значению, но и по особенностям её географического распространения. Пищевыми объектами морских сельдевых рыб могут служить как рыбы (кильки, бычки), так и планктонные организмы.
Многие виды беспозвоночных, являющихся промежуточными хозяевами гельминтов, восприимчивы к условиям среды обитания. Трофический статус водоёма, определяющий разнообразие и продуктивность гидрофауны, является важным фактором в формировании паразитофауны рыб [16]. Паразитические организмы могут регулировать численность своих хозяев [20]. В паразитарных системах патогенность паразита выступает как результат снижения восприимчивости хозяина и связана с нарушением ряда функций его организма: иммунной защиты, стресса, недостатка питания и др. Скрытое течение инвазии переходит в эпизоотию при сочетании высокой численности паразитов с неблагоприятными условиями среды [19]. Наличие и обилие паразитических организмов у рыб может отражать общее благополучие гидробиоценоза, поэтому изучение паразито- и микроценозов морских мигрирующих сельдей – перспективного объекта промысла, является актуальным направлением экологического мониторинга.
Целью настоящей работы было изучение паразитоценоза морских мигрирующих сельдей.
Материалы и методы
С целью изучения паразитофауны морских сельдевых рыб в весенний период 2014‒2015 гг. патологоанатомическому вскрытию было подвергнуто 88 экз. долгинской сельди и 36 экз. каспийского пузанка, выловленных на акватории Северного Каспия. Весной 2015 г. для микробиологических исследований было отобрано по 15 экз. каспийского и большеглазого пузанков и 15 экз. большеглазого пузанка.
При исследованиях рассчитывали экстенсивность инвазии (ЭИ), интенсивность инвазии (ИИ) и индекс обилия (ИО).
Среди микробиологических показателей учитывали число мезофильных аэробных и факультативно-анаэробных микроорганизмов (КМАФАнМ), видовой состав бактерий, их факторы патогенности и антибиотикорезистентность.
Паразитологические и микробиологические исследования проводили в соответствии с общепринятыми методиками [2, 13, 15]. Видовую идентификацию выявленных гельминтов осуществляли по «Определителю паразитов пресноводных рыб СССР» [3] и «Определителю паразитов пресноводных рыб фауны СССР» [4] с использованием стереоскопических микроскопов МБС-10 и биологических микроскопов «Микмед-1» и «Олимпус». Видовую идентификацию выделенных бактерий проводили с помощью определителя Берджи [5].
Результаты и обсуждение
В компонентном паразитарном сообществе долгинской сельди, каспийского и большеглазого пузанков Каспийского моря зарегистрировано семь видов: Mazocraes alosae (Monogenoidea), Diplostomum spathaceum (Trematoda), Ergasilus sieboldi (Crustacea), Pseudopentagramma symmetricum (Trematoda), Corynosoma strumosum (Acanthocephala), Contracaecum sp. (Nematoda), Anisakis schupakovi (Nematoda).
Высокие показатели инвазии отмечены кишечной трематодой P. symmetricum и моногенеей M. alosae, поражающими жаберные лепестки. Стопроцентное заражение P. symmetricum выявлено у каспийского пузанка, однако максимальная интенсивность инвазии (6000 экз.), а также наибольшая зараженность моногенетическими сосальщиками отмечена у долгинской сельди (табл. 1).
Таблица 1
Экстенсивность инвазии паразитами сельдевых видов рыб
в 2014–2015 гг.
|
Вид паразитов |
Локализация |
Экстенсивность инвазии паразитами, % |
|
|
долгинской сельди |
каспийского пузанка |
||
|
Monogenoidea |
|||
|
M. alosae |
Жабры |
91,25 |
46,65 |
|
Crustacea |
|||
|
E. sieboldi |
Жабры |
‒ |
3,35 |
|
Trematoda |
|||
|
D. spathaceum |
Стекловидное тело |
3,85 |
3,35 |
|
P. symmetricum |
Кишечник |
95,10 |
100 |
|
Nematoda |
|||
|
A. schupakovi |
Серозная оболочка кишечника, брыжейка |
62,45 |
11,40 |
|
Contracaecum sp. |
Кишечник |
59,20 |
34,75 |
|
Acanthocephala |
|||
|
C. strumosum |
Наружные стенки кишечника и печени |
7,25 |
13,80 |
Несмотря на высокую численность этих паразитов, выраженных патологических изменений в местах локализации не обнаружено. В меньшей степени трематодами был инвазирован большеглазый пузанок (ЭИ 46,67 %).
В отличие от моногеней, обладающих видоспецифичностью к морским сельдям, кишечные трематоды зарегистрированы у всех каспийских сельдевых видов рыб (в том числе каспийских килек), преимущественно пелагических планктофагов. Такую гостальную приуроченность можно объяснить только особенностями жизненного цикла трематоды, которые должны включать в качестве дополнительного хозяина планктонных беспозвоночных [11]. В нашем случае численное преобладание данных гельминтов у хищной долгинской сельди, вероятно, связано с высоким содержанием инвазированных кормовых объектов в её рационе за счет смешанного питания.
В состав эпидемиологически значимых паразитов сельдевых видов рыб входили круглые черви A. schupakovi, Contracaecum sp. и скребни C. strumosum. Превалентность нематод сем. Anisakidae изменялась в широком диапазоне (от 9,50 до 91,49 %) у различных видов рыб р. Alosa. Так, наибольшее число зараженных особей отмечали у долгинской сельди вследствие более широкого спектра питания. Акантоцефалы в большей степени инвазировали стенки желудочно-кишечного тракта и печень каспийского пузанка (табл. 1). Большеглазый пузанок был свободен от инвазии скребнем. Доля опасных для человека гельминтов в паразитоценозе морских сельдей в среднем не превышала 2,30 %, в то время как трематоды составляли 94,34 %.
Широко распространенные в Волго-Каспийском регионе опасные для здоровья теплокровных животных и человека гельминты A. schupakovi, Contracaecum sp. и C. strumosum паразитируют у сельдевых рыб на стадии личинки [12]. В жизненном цикле контрацекума, характеризующегося широкой специфичностью к хозяевам I и II порядков, участвуют эвригалинные зоопланктонные организмы, поэтому возможность заражения ими различных представителей ихтиофауны велика. Развитие анизакиса и коринозомы, чувствительных к солености воды, протекает при участии только морских форм планктонных животных, что объясняет высокие показатели зараженности у солоноводных видов рыб. Как известно, биология коринозом выделяется среди других скребней широким распространением в жизненных циклах практически всех их представителей паратенического (резервуарного) паразитизма, когда закончившие развитие в промежуточных хозяевах личинки скребней (цистаканты) аккумулируются (без какого-либо дальнейшего развития) и переживают в полости тела и внутренних органах различных видов морских, проходных и даже пресноводных рыб. Тем самым оптимизируется передача личинок паразитов дефинитивным хозяевам [1].
На акватории Северного Каспия природный очаг анизакиозной и коринозомозной инвазии активно поддерживается каспийским тюленем, являющимся окончательным хозяином в паразитарной системе этих эпидемически значимых гельминтов.
Эпизоотическую значимость для своих хозяев представляют трематоды D. spathaceum и ракообразные E. sieboldi. Экстенсивность инвазии и численность D. spathaceum была невысокой (ЭИ 3,85 и 3,35 % у долгинской сельди и каспийского пузанка; ИО 0,12 и 0,13 экз./рыбу соответственно). Заражения вышеперечисленными паразитами у большеглазого пузанка не зарегистрировано. Данный вид диплостоматид является этиологической причиной заболеваний карповых рыб паразитарной катарактой глаз. Поскольку трематоды специфичны к первому промежуточному хозяину – моллюскам рода Lymnaea, то численность паразитов будет напрямую зависеть от числа этой группы беспозвоночных в спектре питания рыб.
Основное заражение трематодами происходит в хорошо прогреваемых акваториях с развитой зарослевой литоралью, обилием моллюсков и рыбоядных птиц. Поступление биогенов приводит к зарастанию литоральной зоны озера и увеличению численности моллюсков – первых промежуточных хозяев диплостомид [18]. Широкое распространение этих видов трематод среди представителей ихтиофауны Волго-Каспийского бассейна доказывает сохранение исторически сложившихся трофических связей между хозяевами в жизненном цикле этих паразитов.
Если зараженность трематодами сопровождается сменой промежуточных хозяев, то инвазия рачками E. sieboldi осуществляется непосредственно из внешней среды и зависит от гидролого-гидрохимического режима водоема [6]. При массовом паразитировании они могут вызывать патологические изменения жаберного аппарата, проявляющиеся в нарушении функции кровообращения, целостности лепестков и обильном слизеотделении. Эргазилус был обнаружен только у каспийского пузанка единично с интенсивностью инвазии 5 экз./рыбу.
В отличие от качественного состава паразитов, их количественные показатели в годовом аспекте характеризовались широкой вариабельностью, при этом экстенсивность заражения выявляемыми паразитами у долгинской сельди понижалась, у каспийского пузанка увеличивалась, что обусловлено пищевыми приоритетами и условиями нагула сельдей в вегетативный период.
В настоящее время, по сравнению с результатами паразитологических исследований, проведенных в 2005–2007 гг., из компонентного паразитарного сообщества морских мигрирующих сельдей исчезли ранее отмечаемые эндемики специфичные трематоды Bunocotyle cingulate. Цикл развития кишечной трематоды протекает при участии веслоногих рачков Limnocalanus и Eurytemora [4], которые являлись традиционными кормовыми объектами сельдевых рыб, но в последние годы либо отсутствуют, либо встречаются единично в их рационе. В таких случаях рыба отдает предпочтение более доступным кормовым объектам, что нарушает жизненный цикл этих гельминтов. Однообразие пищевых компонентов приводит не только к обеднению паразитофауны, но и вызывает большое сходство ее у различных видов рыб [18].
В целом, основу фауны паразитов сельдевых видов рыб формировали специфичные виды гельминтов с прямым и сложным циклами развития, которые сосуществовали в организме своих хозяев на уровне бессимптомного паразитоносительства. Более широким видовым разнообразием паразитов характеризовался каспийский пузанок.
Выявление индикаторных паразитов (D. spathaceum и E. sieboldi), ранее не регистрируемых у морских сельдей, указывает на процессы эвтрофикации водоема [9].
Другим важным показателем состояния экосистемы северной части Каспийского моря является микробиота, ассоциированная с гидробионтами, среди которой наиболее значимой частью являются условно-патогенные бактерии, способные вызывать инфекционные процессы у рыб.
В результате микробиологических исследований каспийского и большеглазого пузанков установлено, что все изученные органы (кишечник, мышцы, печень, жабры) были заражены микроорганизмами (табл. 2).
Таблица 2
Общая микробная обсемененность внутренних органов сельдей Северного Каспия
|
Орган, ткань |
Число микроорганизмов (КОЕ/г), выявленных у пузанка |
|
|
каспийского |
большеглазого |
|
|
Кровь |
Стерильно |
Стерильно |
|
Жабры |
7,4 × 103 |
2,6 × 103 |
|
Мышцы |
2,7 × 102 |
3,8 × 102 |
|
Кишечник |
5,8 × 104 |
6,3 × 104 |
|
Печень |
1,7 × 102 |
1,9 × 102 |
Максимальная численность бактерий выявлена в кишечнике рыб и жабрах (5,8 × 104; 6,3 × 104 и 7,4 × 103; 2,6 × 103 КОЕ/г), что обусловлено функцией пищеварительной системы [7] и непосредственным контактом жаберного аппарата с внешней средой [14].
В микробиоценозе жабр отмечено преобладание энтеробактерий (6,30–11,10 % выделенных микроорганизмов), что связано с доминированием данной группы микроорганизмов в составе бактериопланктона Северного Каспия.
В микроценозе кишечника также преобладали представители сем. Enterobacteriaceae (12,50–22,30 % выделенных микроорганизмов), однако, в отличие от жабр, энтеробактерии кишечника имели автохтонное происхождение и участвовали в пищеварении и обеспечении местного иммунитета.
В мышцах и печени рыб зарегистрированы грамположительные микроорганизмы, энтеробактерии, алкалигенесы и виды с неясным систематическим положением (3,10–12,50 % выделенных микроорганизмов). Поскольку в норме данные органы у рыб стерильны, их бактериальная контаминация могла произойти в результате эндогенного обсеменения, что подтверждалось практически полной идентичностью видового разнообразия микроорганизмов мышц, печени и кишечника.
Эпизоотическая значимость условно-патогенной микрофлоры каспийского и большеглазого пузанков подтверждается наличием факторов патогенности и антибиотикорезистентности бактерий. Один из важных факторов патогенности — это экзоферменты бактерий, с помощью которых последние способны проникать в ткани рыб и вызывать развитие в рыбе патологических процессов [17]. В бактериоценозе сельдей отмечены микроорганизмы, продуцирующие протеазу, лецитиназу, ДНК-азу, гемолизины. Помимо вирулентных свойств, выделенные бактерии проявляли устойчивость к действию антимикробных препаратов. Максимальные показатели резистентности отмечены для бензилпенициллина и ампинициллина (100 % устойчивых изолятов), а минимальные – для офлоксацина (27,5 % устойчивых изолятов).
Однако, микробиоценоз морских сельдей был представлен не только сапрофитными и условно-патогенными бактериями, но и микроскопическими грибами, гифы которых были определены при микроскопировании мазков отпечатков печени и селезенки этих рыб. Подобные структурные элементы микромицетов ранее были выявлены во внутренних органах каспийских килек. При этом у сельдей отмечали латентно протекающую микоинфекцию, а у каспийских килек висцеральное микотическое поражение сочеталось с опухолеобразованием [8].
В результате обследования весной 2015 г. в печени и селезенке у 53,4 % долгинской сельди, 66,7 % каспийского пузанка и 20,0 % большеглазого пузанка выявлены гифы микромицетов. По окраске и структуре гифов грибов полученные препараты схожи с мазками-отпечатками каспийских килек (рис.).
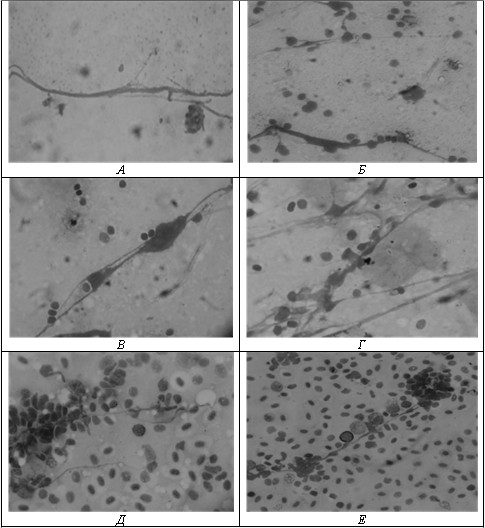
Рис. Гифы микроскопических грибов в печени и селезенке долгинской сельди (А, Б), каспийского пузанка (В, Г), большеглазого пузанка (Д, Е) (окраска по Романовскому‒Гимзе, ув. × 1000)
При визуальном осмотре внутренних органов сельдей патологий и новообразований как у каспийских килек, не регистрировали. Однако, отсутствие макроскопических узелков (гранулем, которые, как правило, появляются в ответ на грибковую инфекцию) не исключает висцерального микозного поражения, поэтому проводимое микроскопирование мазков тканей печени и селезенки позволяет обнаружить в этих органах патогенного агента на начальных стадиях инфицирования.
Заключение
Компонентное паразитарное сообщество морских мигрирующих сельдей было представлено морскими и эвригалинными видами паразитов с моноксенным и поликсенным циклами развития. Качественный состав паразитов этих рыб характеризовался низким видовым разнообразием, количественные показатели – широкой вариабельностью. При этом на видовую изменчивость паразитов морских мигрирующих сельдей оказывали влияние пищевые приоритеты гидробионтов, на динамику инвазированных особей ‒ условия среды обитания, биологические особенности паразитов и их хозяев. Относительно постоянный уровень инвазии рыб эпизоотически и эпидемиологически значимыми паразитами свидетельствует об устойчивости паразито-хозяинных отношений внутри паразитарной системы сельдевых видов рыб.
Широко распространенные на акватории Каспийского моря сапрофитные и условно-патогенные микроорганизмы контаминировали внутренние органы гидробионтов на уровне бактерионосительства, не провоцируя развитие инфекционного процесса. Наличие гифов микроскопических грибов в органах долгинской сельди, каспийского пузанка и большеглазого пузанка, которые не вызывали выраженных клинических проявлений, может указывать на некоторое снижение резистентности организма сельдей в нерестовый период.
Литература
1. Атрашкевич Г. И. Скребни рода Corynosoma Lune, 1904 (Acanthocephales; Polymorphidae) в Охотском море и паразитарная система доминирующего вида C. strumosum (Rudolphi, 1819) // Паразитология в XXI веке – проблемы, методы, решения: Матер. IV Всерос. Съезда Параз. О-ва при РАН (Санкт-Петербург, 20‒25 октября 2008 г.). В 2-х т. – СПб: Лема, 2008. – Т 1. – С. 38–42.
2. Быховская–Павловская И. Е. Паразиты рыб: Руководство по изучению. – Ленинград: Наука, 1985. – 121 с.
3. Быховская–Павловская И. Е., Гусев А. В., Дубинина М. Н. и др. Определитель паразитов пресноводных рыб СССР. – Москва–Ленинград: Академия наук СССР, 1962. – 776 с.
4. Бауэр О. Н. Определитель паразитов пресноводных рыб фауны СССР. – Л.: Наука, 1987. – Т. 3. – 583 с.
5. Хоулт Дж., Криг Н. Определитель бактерий Берджи. В 2-х. т. – М.: Изд-во Мир, 1997. – 799 с.
6. Бурдуковская Т. Г. Ракообразные (Crustacea) – паразиты рыб озера Байкал: гостально-пространственное распределение и экология: дис. ... канд. биол. наук. – Улан-Удэ, 2006. – 121 с.
7. Воробьев А. А. Медицинская микробиология, вирусология и иммунология. – М.: МИА, 2004. – 690 с.
8. Воронина Е. А., Дубовская А. В. Висцеральные опухолеобразования у анчоусовидной кильки (Clupeonella engrauliformis) Каспийского моря // Рыбное хозяйство. – 2011. – № 4. – С. 72–74.
9. Заостровцева С. К. Анализ паразитофауны рыб Вислинского залива (Балтийское море) // Биология внутренних вод. – 2009. – № 4. – С. 87–92.
10. Калмыков В. А., Ходоревская Р. П., Абдусамадов А. С., Смирнов А. В. Обзор развития прибрежного рыболовства морских сельдей закидными неводами на западном побережье Каспийского моря (Российский регион) // Вопросы рыболовства. – 2012. – Т. 13, № 4 (52). – С. 773–778.
11. Корнейчук Ю. М., Барзегар М. Трематода Pronoprymna ventricosa (Rud., 1819) – паразит каспийских сельдей // Морской экологический журнал. – 2005. – № 1. – С. 45–47.
12. Ларцева Л. В., Проскурина В. В. Состояние паразитофауны и микрофлоры гидробионтов Волго-Каспийского региона на рубеже XXI века. – Астрахань: КаспНИРХ, 2003. – 80 с.
13. Мусселиус В. А., Ванятинский В. Ф., Вихман А. А. Лабораторный практикум по болезням рыб. – М.: Легкая и пищевая пром-ть, 1983. – 296 с.
14. Морозова М. А., Ларцева Л. В. Микробные сообщества гидроэкосистемы Нижнего Дона и Таганского залива // Естественные науки. – 2012. – № 2 (39). – С. 50–56.
15. Нетрусов А. И., Егорова М. А., Захарчук Л. М. Практикум по микробиологии: учеб. пособие для студ. высш. учеб. заведений. – М.: Академия, 2005. – 608 с.
16. Новохацкая О. В. Паразитофауна рыб эвтрофируемых озер (на примере Сямозера): автореф. дис. … канд. биол. наук. – Спб, 2008. – 26 с.
17. Обухова О. В., Ларцева Л. В. Микробиоценоз воды и судака (Sander lucioperca) в дельте р. Волги: монография. – СПб: Проспект науки, 2015. – 224 с.
18. Румянцев Е. А. Влияние некоторых факторов на паразитофауну рыб при интродукции в озера Карелии // Паразитология. – 1975. – IX, № 4. – С. 305–311.
19. Трухачев В. И., Толоконников В. П., Лысенко И. О. Научные основы экологической паразитологии. – Ставрополь: АГРУС, 2005. – 414 с.
20. Шабунов А. А., Радченко Н. М. Паразиты рыб, земноводных и чайковых птиц в экосистемах крупных водоемов Вологодской области: монография. – Вологда: ВоГТУ, 2012. – 243 с.
© 2017 The Authors. Published by All-Russian Scientific Research Institute of Fundamental and Applied Parasitology of Animals and Plants named after K.I. Skryabin. This is an open access article under the Agreement of 02.07.2014 (Russian Science Citation Index (RSCI)http://elibrary.ru/projects/citation/cit_index.asp) and the Agreement of 12.06.2014 (CABI.org / Human Sciences section: http://www.cabi.org/Uploads/CABI/publishing/fulltext-products/cabi-fulltext-material-from-journals-b...